TranslatorПредупреждение Меню сайтаФорма входаСтатистикаCopyright © Далгатов, 2011-2016 Все права защищены
 
 Онлайн всего: 1 Гостей: 1 Пользователей: 0 Поиск |
Коган А. Б., Основы физиологии высшей нервной деятельности: Учеб. для биол. спец. вузов.- 2-е изд., перераб. и доп. - М.: Высш. шк., 1988. - 368 с.: ил. ISBN 5-06-001444-4.
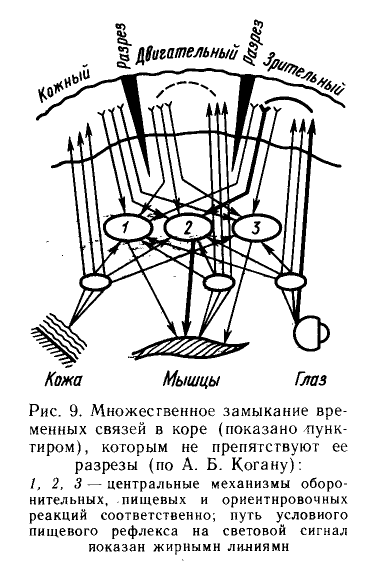
Весьма серьёзное непонимание профессором А. Б. Коганом сути происходящего на рис. 9. На рисунке цифрами 1 и 2 обозначены оборонительный и пищевой мотивационные центры гипоталамуса. Цифрой 3 обозначена ретикулярная формация ствола мозга, "ответственная" за ориентировочную реакцию. Без этих структур невозможно замыкание временных связей в коре. Кору профессор А. Б. Коган разрезал аккуратно вдоль границ анализаторов. Что мы реально наблюдаем? Нет, не жирные линии. Там есть линии тоньше. Смотрим. Глаз - маленький овал - большой овал с цифрой 3 - мышцы. Цифра 3 указывает на ретикулярную формацию. Никакой коры. Этот путь наиболее оптимален. Меньше переключений. То есть, у пищевого рефлекса два пути в этой редакции. Но: "Специальное внимание было уделено также выяснению участия гипоталамуса в условнорефлекторном процессе... Установлено также, что коротколатентный ответ (20 мс и менее) на звук регистрировался до обучения у многих нейронов гипоталамуса. При выработанном условном пищевом рефлексе ответы на сигнальный раздражитель имели скрытый период до 40-80 мс, предшествуя поведенческому ответу, развивающемуся через 80-120 мс после включения сигнального раздражителя. Ответ нейронов в последовательности сочетаний появлялся также раньше поведенческой реакции. Эти данные интерпретируются как доказательство участия гипоталамуса в интеграции информации о свойствах подкрепления для осуществления конечного адаптивного эффекта. Такой вывод основывается главным образом на том, что ответ на условный сигнал возникает в последовательности сочетаний раньше, чем в других областях мозга, включая кору" (Б. И. Котляр, "Пластичность нервной системы", М.; Изд-во МГУ, 1986, стр. 85). Выше указан звук, как сигнальный раздражитель. Принципиальной разницы нет. Рисунок не соответствует действительности. Смотрим, как "движется" жирная линия. Отойдя от глаза, она один раз переключается (верхние бугорки четверохолмия?). Далее, поднимается в зрительный анализатор. Переключившись в зрительном анализаторе, она опускается к большому овалу с цифрой 2. Это "центральный механизм пищевой реакции", как пишет профессор Коган. Видимо, пищевой центр гипоталамуса. От него жирная стрелка уходит вниз к мышце. Такого не существует. Такого пути нет. Если речь о ретикуло-спинальном тракте, то линия должна была двинуться к овалу с цифрой 3. Может это ошибка и вместо слова "мышцы" должно было стоять слово "железа"? Кроме того, жирные линии почему-то игнорируют двигательный анализатор коры. Зрительный анализатор не взаимодействует с двигательным. "Множественное замыкание" будет произведено только с "санкции" ретикулярной формации. Даже у спинального препарата кошки (ниже). "...Ко второй группе отнесена постсинаптическая модуляция автономной ритмики пейсмекерных клеток, широко встречающихся в нервной системе беспозвоночных" (Котляр, 1986). Пейсмекерность вышеуказанных клеток указывает на их родственность пейсмекерным ретикулярным нейронам позвоночных. Почему акцентирование внимание на ретикулярной формации, как это делает Пенфилд, недостаточно? Потому, что без выделения особой роли гипоталамуса, полноценного объяснения не достичь (в гипоталамусе ретикулярная формация сама занимает значительный объём). Вышеуказанный опыт профессора Когана подтверждает невозможность автономного замыкания корково-корковых временных связей. Хотя он пытается доказывать обратное. Я пишу о связях между корковыми анализаторами. Новая кора самостоятельно "не думает", то есть не замыкает ассоциативных связей. Принципиально необходимо участие HR-системы (гипоталамо-ретикулярной). Гипоталамус необходим для классических условных рефлексов. Для индифферентных раздражителей достаточно и одной ретикулярной формации. "В школе И. П. Павлова одновременно с осуществлением обширной программы исследований слюнных условных рефлексов было начато физиологическое изучение особой формы временной связи - ассоциаций между так называемыми индифферентными раздражителями (Панферов, 1926; Зелёный и др., 1928; Шохор, 1929; Подкопаев, 1934; Нарбутович, Подкопаев, 1936). Для формирования этого типа временных связей, в отличие от классических условных рефлексов, образуемых на основе биологически значимой мотивации (пищевой, половой, защитной), оказалось достаточным участия лишь возбуждения, связанного с ориентировочной реакцией" (Б. И. Котляр, 1986). Что по поводу индифферентных раздражителей? "Исследование соотношения физиологических и психических показателей мышления, а также восприятия различных индифферентных и значимых сигналов, в том числе осознаваемых и неосознаваемых словесных раздражителей, позволило приступить к изучению физиологических механизмов психических проявлений" (Котляр, 1986). "Индифферентные и значимые сигналы, в том числе осознаваемые и неосознаваемые словесные раздражители...". Что это? Ваш ассоциативный процесс... Визуальные представления там же. Любая помысленная вами ассоциация (любая мысль) предварительно замыкается ретикулярной формацией ствола мозга. Мыслит HR-система (гипоталамо-ретикулярная). Замыкая колоночные нейронные структуры в ассоциативных полях новой коры. Это работа ассоциативного процесса в норме. Из неё прямо вытекают и ассоциативный дефект при шизофрении и скачка идей (fuga idearum) при мании. Поиск и нахождение конкретных энграмм, происходит при помощи фронтальных ассоциативных полей. Ассоциативный процесс имеет две составляющие. Первая - тоническая составляющая (Котляр). Вторая - фазическая составляющая, сама временная связь групп нейронов. Мысль есть вторая составляющая. При шизофрении она не нарушена (но нарушено "удержание" её на надлежащих путях, об этом ниже). Шизофреник может прекрасно разобраться в философской статье, как писал Блёйлер. В чём же проблема? Во-первых, в тонической составляющей. Она "порождается" ретикулярной формацией. Её дисфункция, в выраженных случаях, приводит к шперрунгу. Во-вторых, в удержании ассоциаций на надлежащих путях. Это функция GABAergic neurons неокортекса (см.).
Возможным анатомическим субстратом, указанных двух составляющих, являются: для первой - норадренергическое голубоватое ядро (locus coeruleus, отдел ретикулярной формации) ствола мозга (единственный, как предполагается, источник норадреналина в коре больших полушарий), для второй - холинергическое базальное крупноклеточное ядро переднего мозга, или ядро Мейнерта. Подробно: www.totalpsychiatry.org Необходимо добавить, что "мощность гамма-ритма отражает активацию процессов обработки информации в коре больших полушарий" и "...генерация гамма-ритма критически зависит от холинергической иннервации, поступающей в кору больших полушарий из БКЯ" (Чернышев Б. В., Тимофеева Н. О., Мацелепа О. Б., Семикопная И., 2011, БКЯ - базальное крупноклеточное ядро Мейнерта).
Тоническая составляющая - ассоциация (мысль, представление) - тоническая составляющая - ассоциация (мысль, представление). Вот он, ассоциативный процесс Homo sapiens.
"...Отсюда, по мнению автора, следует, что функции коры не могут проявиться только при раздражении коры. Все функции коры проявляются только после того, как они получат дополнительно естественный импульс, идущий из другого пункта мозга". dommedika.com 1. "Обобщая итоги многолетних исследований, П. С. Купалов (1963) выделил два механизма, участвующих в условнорефлекторной деятельности: первый, настроечный, регулирующий общий тонус и создающий определённый уровень возбудимости и работоспособности нервных центров, второй, запускающий процессы, которые осуществляют ту или иную условную реакцию".
7. "И. П. Павлов придавал большое значение исследованиям Л. А. Орбели и высоко их оценивал. Так, представляя Орбели в действительные члены Академии наук, он отмечал: "Его главная заслуга, высоко ставящая его среди современных физиологов, есть установка факта прямого влияния симпатических волокон на скелетную мускулатуру и центральную нервную систему, факта, решающего почти столетнюю загадку о так называемой трофической иннервации, которая должна обнять огромнейшую массу как физиологических, так и патологических явлений животного организма" (В. Ф. Ананин, 1992). 8. "Автор концепции об адаптационной функции симпатической нервной системы Л. А. Орбели не ограничивал её рамками периферических органов, в том числе и скелетной мускулатуры. Он считал, что и сама центральная нервная система испытывает адаптационное влияние со стороны симпатической нервной системы. Так, Орбели писал: "Можно считать вполне установленным, что симпатическая нервная система (а в некоторых случаях и парасимпатическая!) оказывает прямое регулирующее влияние на все возбудимые ткани, в силу чего высшие отделы мозга могут через посредство симпатической системы создавать во всех частях рефлекторного аппарата адаптационные влияния в широком смысле этого слова, то есть известную, соответствующую условиям времени предуготовленность рефлекторного аппарата к выполнению его функций" (Л. А. Орбели, 1926). Однако конкретные механизмы этой адаптации так и остались не раскрытыми" (В. Ф. Ананин, 1992). Объясню последнюю цитату. Академик Орбели в 1926 году прямо предвосхищает то, что в 1958 году указал Пенфилд. По академику Орбели, получается, что симпатическая система, а именно ретикулярная формация, прямо регулирует высшие отделы мозга. По Пенфилду, высшие отделы мозга, neocortex, могут осуществлять свои функции только при получении дополнительного импульса из ретикулярной формации. По профессору Котляру, любая ассоциация в кортикальных структурах возможна только при участии тонической составляющей ассоциативного процесса. Эту тоническую составляющую генерирует ретикулярная формация. Орбели - Пенфилд - Котляр.
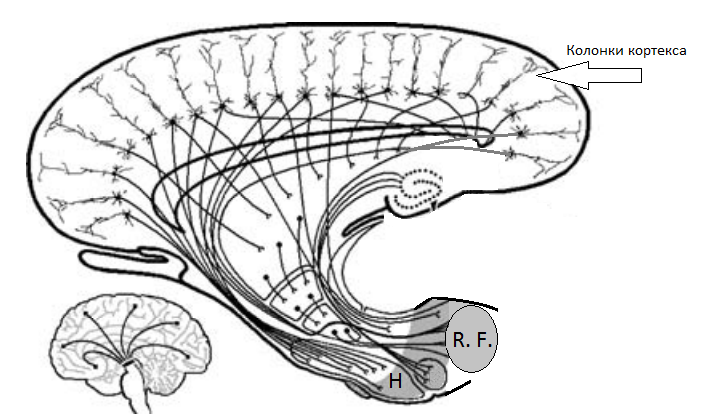
H - Hypothalamus, R. F.- Reticular Formation Определение ассоциативного процесса, по сути, уже здесь: "Иррадиация (от лат. irradiare - сиять, испускать лучи) - способность нервного процесса распространяться из места своего возникновения на другие нервные элементы. Согласно И. П. Павлову, иррадиация возбуждения лежит в основе генерализации условного рефлекса и зависит от интенсивности стимула. В настоящее время установлено, что эффект генерализации является результатом включения активирующих подкорковых структур (см. Мозг)..." ("Психологический словарь", под общ. ред. А. В. Петровского, М. Г. Ярошевского.- М.: Политиздат, 1985.- 431 с., стр. 128). В свете вышесказанного, интерпретация указанного опыта профессором А. Б. Коганом выглядит совершенным нонсенсом. Оно антинаучно в свете следующего: "К началу текущего столетия, - писал Н. Г. Колосов (1974), - было накоплено большое количество убедительных фактов, доказывающих рефлекторную фунцкцию вегетативных ганглиев и позволяющих рассматривать эти последние как настоящие перефирические рефлекторные центры, способные осуществлять свою замыкательную функцию и без участия ЦНС" (В. Ф. Ананин, Проблемы неврологии ХХ века: Крушение иллюзий и новые открытия: монография. М.: изд-во РУДН и Биомединформ, 1992. 120 с., стр. 16). Ещё цитата: "Подводя итоги исследований своего коллектива, И. А. Булыгин (1981) писал: "Вегетативные ганглии выполняют рефлекторную функцию, в основе которой лежит синаптическое соединение в ганглиях афферентных, эфферентных и вставочных вегетативных нейронов. Тела афферентных нейронов, соединяющих внутренние органы с экстрамуральными симпатическими ганглиями и ЦНС, лежат в интра- и экстрамуральных ганглиях и по ходу вегетативных нервов. Они принимают участие в образовании афферентного звена истинных периферических рефлексов, замыкающихся в превертебральных и паравертебральных симпатических ганглиях и на уровне спинного мозга" (там же, стр. 17). |